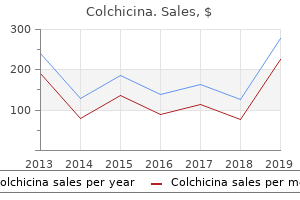
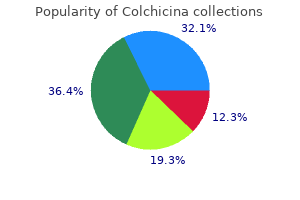
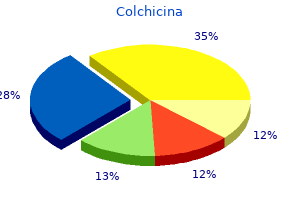
Colchicina"Order colchicina 0.5 mg with visa, antibiotics for uti and bv". By: C. Chris, MD Co-Director, Florida State University College of Medicine Cannabinoids are efficacious in suppressing nociception in preclinical models of neuropathic pain induced by diabetes infection zombie book buy generic colchicina 0.5mg. Endocannabinoid modulators also show anti-hyperalgesic efficacy in models of neuropathic pain. Recent literature also suggests that the endocannabinoid signaling system can be targeted to suppress both the evolution and progression of cancer. Changes in endocannabinoid signaling and the activity of endocannabinoid hydrolyzing enzymes have also been demonstrated in bone cancer models. Finally, in vitro studies suggest multiple antitumor effects of cannabinoids (inhibition of cell proliferation and migration, induction of apoptosis) that may contribute to a highly favorable therapeutic profile of the in vivo efficacy of cannabinoids. More work is necessary to determine whether cannabinoids exhibit sufficiently high therapeutic potential to reduce pain, promote antitumor effects, and improve quality of life in cancer patients. There is little substantial evidence of cannabinoid analgesia for postoperative or non-neuropathic chronic pain. More work is necessary to determine whether adjunctive treatment with cannabinoids enhances the efficacy and reduces the side effects of other analgesics. In four of these studies, pain relief was greater in patients randomized to cannabinoid than to placebo. Another more recently published study suggested a modest degree of analgesic efficacy when two concentrations of smoked cannabis were compared with a placebo preparation in subjects with a range of central and peripheral neuropathic pain conditions, mainly complex regional pain syndrome type 1 (Wilsey et al 2008). Using internationally accepted criteria for defining the severity of adverse events, the authors reported that although "non-serious" acute adverse events were associated with cannabinoid treatment (rate ratio, 1. The authors drew attention to the paucity of safety data related to the effects of long-term therapeutic exposure to cannabinoids. Finally, implications for the development of cannabinoids as therapeutics do need to take into account the impact of cannabis intoxication on the cognitive skills required for driving motor vehicles (Asbridge et al 2012). Implications of the Risk for Mental Illness with Cannabis Abuse In any discussion of the evidence of the clinical potential of cannabinoids, the issue of potential long-term mental health risks associated with therapeutic cannabinoid administration requires serious consideration. There is a now a substantial and consistent epidemiological literature supporting a dose-related association between cannabis abuse and subsequent long-term risk for the development of psychotic illness or schizophrenia (see Rice 2008a, 2008b). A systematic review encompassing nine studies consistently identified such findings in studies with different methodology in different settings (Semple et al 2005). Cannabis users are two to three times more likely for serious psychotic illness to subsequently develop, including schizophrenia, than non-cannabis users are. Similar findings were reported in a subsequent systematic review (Moore et al 2010). This observation is strengthened by a clear dose-response relationship between cannabis use and schizophrenia or the development of psychotic symptoms (Zammit et al 2002, Henquet et al 2005, Di Forti et al 2009). There is also a genetic element inherent in such risk: individuals who carry a functional polymorphism in the gene encoding the enzyme catechol O-methyltransferase are more likely to exhibit psychotic symptoms and to develop schizophrenia if they use cannabis (Caspi et al 2005). Polymorphisms in this same enzyme are associated with variations in pain sensitivity and risk for the development of chronic pain conditions such as temporomandibular joint dysfunction (Diatchenko et al 2005). It should be noted that many of these data have emerged largely from studies in adolescents, so the extent to which they can be generalized to the wider population and specifically to patients is unknown. Finally, a frequent criticism of such epidemiological studies is the "self-medication" hypothesis whereby individuals with early preclinical psychosis could self-medicate with cannabis-but this hypothesis has recently been refuted (Henquet et al 2005). The therapeutic trials of cannabinoids for neuropathic pain reported to date are insufficiently powered to detect such infrequent but significant adverse events. Furthermore, although some therapeutic trials have monitored patients for up to 1 year, this may be an insufficient period to detect the long-term adverse effects of cannabinoids. It is impossible to disregard the implications of these facts for cannabinoids being developed for chronic regular administration in patients with long-term conditions such as neuropathic pain. However, these data may reveal insight into the neurobiology of depression (Hill and Gorzalka 2009). More work is necessary to determine whether peripherally restricted modulators of the endocannabinoid system would produce a more circumscribed and beneficial spectrum of therapeutic efficacy in humans than brain-penetrant therapeutic agents would. However, the implications of the strong and consistent epidemiological data associating cannabis misuse with a dose-dependent long-term risk for mental illness cannot be isolated from any discussion of the therapeutic use of potent cannabinoids, especially in the context of neuropathic pain, for which regular longterm therapy is required. Krauspe R, Schmidt M, Schaible H-G: Sensory innervation of the anterior cruciate ligament: an electrophysiological study of the response properties of single identified mechanoreceptors in the cat, Journal of Joint and Bone Surgery bacteria 7th grade science discount colchicina 0.5 mg free shipping. LeBars D, Villanueva L: Electrophysiological evidence for the activation of descending inhibitory controls by nociceptive afferent pathways.
Thus, results from microstimulation experiments involving cutaneous C fibers best match the hypothesis that there is a small group of itch-mediating C fibers among a much larger population of pain-mediating units antibiotic resistance gmo buy cheap colchicina 0.5 mg. Indeed, C fibers responding to the application of histamine in parallel with the itch ratings of subjects have been discovered among the group of mechano-insensitive C afferents (Schmelz et al 1997), thus suggesting that there is a specific pathway for itch. Hence, this fiber type cannot account for the prolonged itch induced by the intradermal application of histamine. In line with the large innervation territories of these fibers, two-point discrimination for histamine-induced itch is poor (15 cm in the upper part of the arm) (Wahlgren and Ekblom 1996). The excellent locognosia for histamine-induced itch in the hand (Koltzenburg et al 1993) might therefore be based on central processing compensating for low spatial resolution in the periphery. The relative prevalence of the different C-fiber types in human skin nerves has been estimated from recordings in the superficial peroneal nerve (Schmidt et al 1997). Polymodal nociceptors, which respond to mechanical, heat, and chemical stimuli, are about four times as abundant as mechanoinsensitive nociceptors in young healthy volunteers, but their proportion decreases in the elderly (2. Among the mechano-insensitive afferent C fibers is a subset of units that have a strong and sustained response to histamine. Specific activation of histamine-positive chemo-nociceptors by prostaglandin E2 (Schmelz et al 2003) in combination with the pruritogenic effects of prostaglandins (Neisius et al 2002) provides a strong argument for a specific neuronal system for itch processing that is separate from the pain pathway (Ikoma et al 2006). Absence of an axon reflex flare therefore suggests that the itch is independent of histaminesensitive C fibers. Itch without an axon reflex flare can also be elicited by weak electrical stimulation (Shelley and Arthur 1957, Ikoma et al 2005), further evidence that the sensation of itch can be dissociated from cutaneous vasodilatation. Cowhage spicules inserted into human skin produce itch at an intensity comparable to that following the application of histamine (LaMotte et al 2009, Sikand et al 2009). However, mechano-responsive "polymodal" C-fiber afferents, the most common type of afferent C fibers in human skin (Schmidt et al 1995), can be activated by cowhage in the cat (Tuckett and Wei 1987) and, according to recent studies, also in nonhuman primates (Johanek et al 2007, 2008) and in human volunteers (Namer et al 2008). These fibers are unresponsive to histamine and not involved in sustained axon reflex flare reactions (Schmelz et al 2000b). This is consistent with the observation that cowhage-induced itch is not accompanied by a widespread axon reflex flare (Shelley and Arthur 1955, 1957; Johanek et al 2007). Although in humans the segregation between histamine-positive, mechano-insensitive fibers and cowhage-positive mechanosensitive fibers is clearcut, in monkey, mechanosensitive C fibers also responded to histamine (Johanek et al 2008). The different histamine response might be explained by higher histamine concentrations with intradermal injection than with iontophoresis. A fibers responding to the insertion of cowhage for several minutes (Ringkamp et al 2011) suggest an additional role of afferent input from myelinated fibers. Differential block of myelinated afferents does not reduce capsaicin-induced pain and only slightly reduces histamine-induced itch; however, it massively reduces cowhage-induced itch, at least in part of the subjects (Ringkamp et al 2011). The exact role of A-fiber input for cowhage-induced itch is unclear because the reduced skin temperature induced by the nerve blocking maneuver in these experiments might also reduce cowhageinduced activation. Given that cowhage spicules can activate a large proportion of polymodal nociceptors, we face a major problem in explaining why activation of these fibers by heat or by scratching actually inhibits itch whereas activation by 40 Act. Conduction latencies of these three marked fibers in response to successive electrical stimulation at the receptive field are plotted from top to bottom. When activated by mechanical (von Frey filament, inactivated cowhage spicules), chemical (active cowhage, histamine), or heat test stimuli, C fibers exhibit an activity-dependent increase in response latency followed by gradual normalization ("marking"). The mechano-responsive fiber (green squares) is activated during mechanical stimulation with the von Frey filament and during the application of inactive cowhage, but lasting activation is seen only after the application of active cowhage. In contrast, mechano-insensitive fibers (purple and salmon triangles) do not respond to cowhage stimulation but are activated following histamine iontophoresis. At the right side of the panel, the itch ratings of the subject are depicted, which were assessed during this experiment. Ratings are given on a numerical rating scale from 0 (0 = no itch) to 10 (10 = maximal imaginable itch).
In neurons with convergent input from skin and muscle, descending inhibition is much stronger for responses to deep input than for responses to cutaneous input (Yu and Mense 1990) antibiotics for acne birth control buy online colchicina. Motor Reflexes Noxious stimulation of joint afferents (Ferrell et al 1988, Schaible and Grubb 1993) and muscle afferents (Mense 1993, 1997) can evoke nociceptive withdrawal reflexes. During acute chemical stimulation of the knee and electrical stimulation of muscle nerves (Woolf and Wall 1986) and during inflammation in the joint (Ferrell et al 1988, He et al 1988), spinal motor reflexes are enhanced. During the development of K/C joint inflammation, progressive inhibition rather than facilitation developed in some -motoneurons (He et al 1988). Experimental myositis decreased reflex activation of -motoneurons, and, in patients, painful muscles exhibit low electromyographic activity (Mense 1997). Such changes may possibly contribute to the muscle atrophy and loss of force often observed during chronic muscle and joint diseases (Mense 1997, Schaible and Grubb 1993). However, muscle spasms may be elicited by painful trigger points in adjacent muscles, articular dysfunction, and ligamentous strain (Mense 1997). The secondary hyperalgesia caused by joint inflammation can last several weeks, and this hypersensitivity is associated with enhanced responses of spinal cord neurons to A and C fiber input (Martindale et al 2007). These changes in the spinal cord are likely to account for the pronounced forms of referred pain in deep tissue that are induced by noxious stimulation of deep tissue in humans (Arendt-Nielsen et al 2000). Responses to noxious compression of the knee increased markedly, and within 1 hour the neuron also started to respond to pressure applied to the ankle and the paw. Because of the development of hyperexcitability, the population of neurons responding to stimulation expands in the segments with input from inflamed deep tissue (Hoheisel et al 1994). Primary afferents from deep tissue (muscle and joint) evoke more prolonged facilitation of a nociceptive flexor reflex than cutaneous afferents do (Woolf and Wall 1986), and injection of capsaicin into deep tissue elicits more prolonged hyperalgesia than does injection of capsaicin into the skin (Sluka 2002). A, Histogram showing the responses of a spinal cord neuron to noxious pressure applied to the knee, ankle, and paw before and after injection of kaolin and carrageenan (K/C) into the knee joint. B, Receptive field (dark area) of the neuron before (control) and during knee joint inflammation (3 hours after K/C injection). C, Area of referred pain in the lower part of the leg of control persons and patients with knee osteoarthritis after local injection of 6% NaCl (a short-lasting painful stimulus) into the tibial muscle near the knee joint. Medullary areas of pain control systems are activated during acute and chronic joint inflammation (Pinto et al 2007). At the acute stage of inflammation, tonic descending inhibition (Cervero et al 1991, Schaible et al 1991, Danziger et al 1999) and heterotopic inhibitory influences (Calvino et al 1987; Danziger et al 1999, 2001) are increased, but this is not sustained at the chronic stage of inflammation (Danziger et al 1999, 2001). On the other hand, descending excitation may support the expansion of receptive fields into healthy areas (Vanegas and Schaible 2004). During acute inflammation this pattern changes in that these mediators are released even when the joint is stimulated at innocuous intensity (Hope et al 1990; Schaible et al 1990, 1994). As an indicator of spinal release of substance P, movements of an arthritic joint were found to induce internalization of the neurokinin 1 receptor (whose expression in the superficial dorsal horn increased during monarthritis) (Sharif Naeini et al 2005). Thus, the milieu in the spinal cord is altered and is likely to change sensory processing. Excitatory neuropeptides facilitate the responses of spinal cord neurons, and they may "open" synaptic pathways such that more neurons respond to stimulation (Mense 1997). These antagonists both attenuated the development of inflammation-evoked hyperexcitability and reduced established hyperexcitability (Neugebauer et al 1995, 1996a, 1996b). However, the neuropeptide receptor antagonists are less antinociceptive than the glutamate receptor antagonists. Topical spinal application of the nonselective Cox inhibitor indomethacin before inflammation attenuated the increase in responses during the development of inflammation, thus showing that spinal prostaglandins further the generation of hyperexcitability. Synaptic Activation and Sensitization of Spinal Cord Neurons with Joint Input Generation and maintenance of central sensitization are produced by the action of transmitter/receptor systems in the spinal cord. During sensitization, primary afferent neurons release more transmitter from their spinal terminations on peripheral stimulation (presynaptic component). Buy generic colchicina 0.5mg. Antimicrobials | Six Classes 2017 videos.
|